|
Antony Research Group Department of Biochemistry and Molecular Biology Saint Louis University School of Medicine |
|

|
Antony Research Group Department of Biochemistry and Molecular Biology Saint Louis University School of Medicine |
|
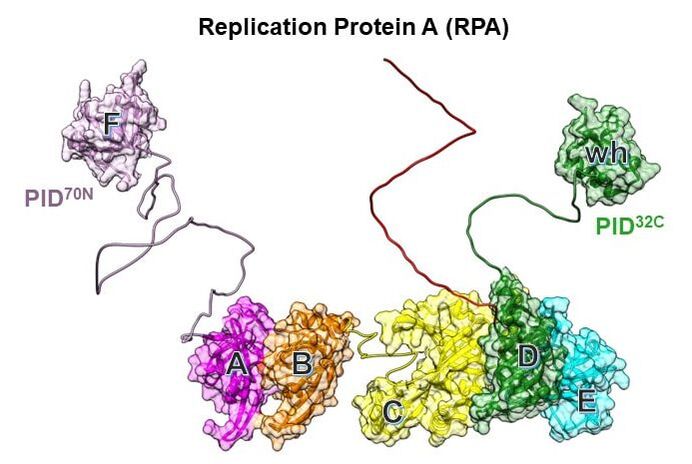
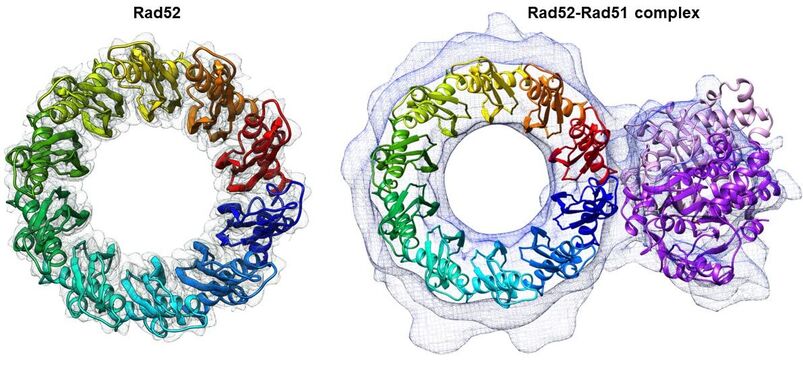
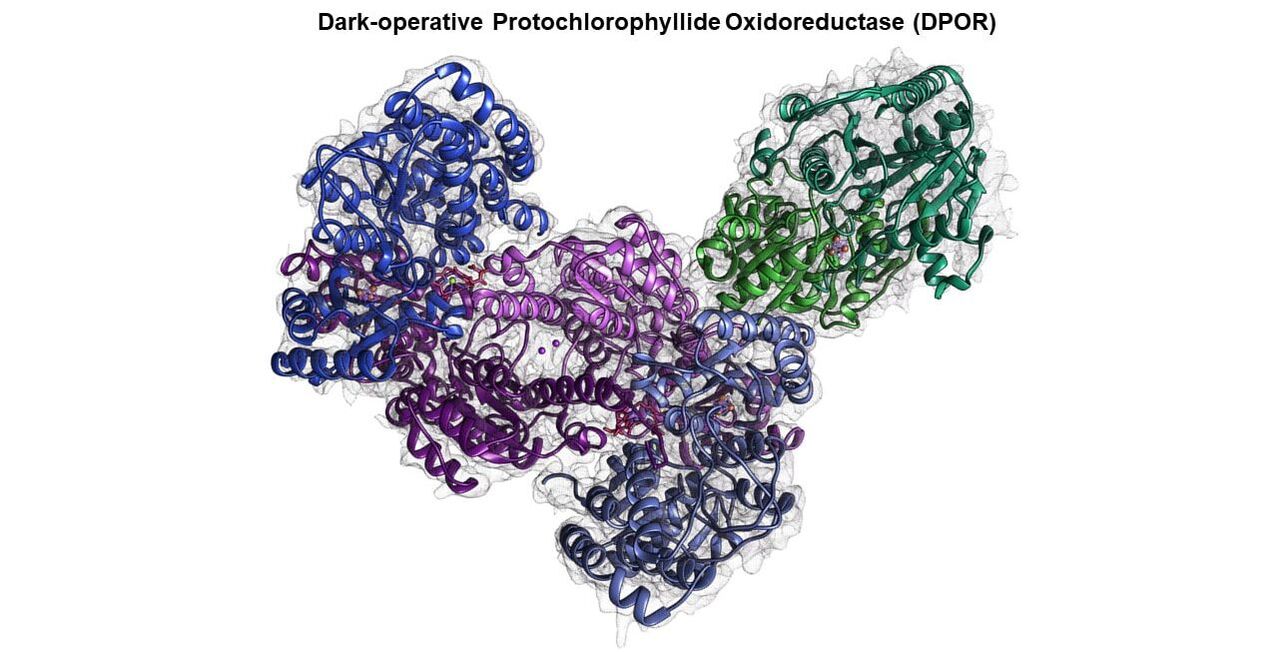
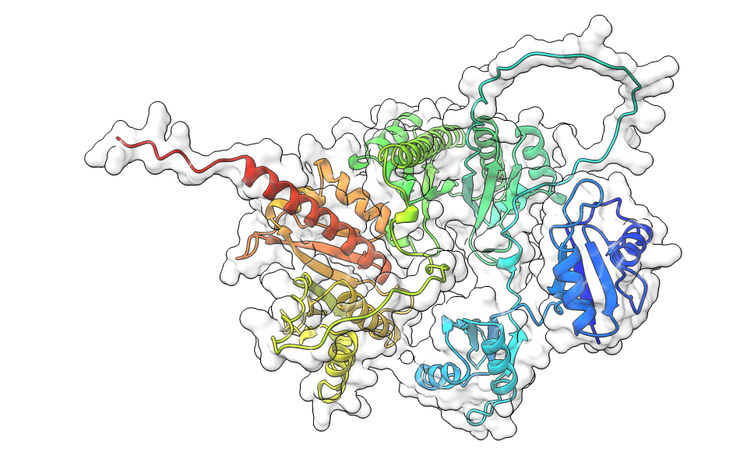

|

|

|
